Université de Liège - Centre de l'Oxygène, Recherche et Développement (CORD)
Université de Liège - Centre de l'Oxygène, Recherche et Développement (CORD)
L'oxygène et la vie: tome 1 - Initiation au métabolisme de l'oxygène
C Deby et G Deby-Dupont
Université de Liège - Centre de l'Oxygène, Recherche et Développement (CORD)
L'oxygène et la vie: tome 1 - Initiation au métabolisme de l'oxygène
C Deby et G Deby-Dupont
Les chapitres précédents ont montré que l'oxygène subit une série de transformations dans les organismes, principalement sous l'action d'enzymes. On peut donc parler d'un véritable métabolisme de l'oxygène, un secteur fort méconnu de la biochimie, mais qui prend de plus en plus d'importance dans la compréhension d'un nombre de plus en plus grand de domaines de la physiologie et de la pathologie de la plupart des êtres vivants.
Néanmoins, il faut avertir le lecteur que la transposition de phénomènes étudiés in vitro à l'échelle in vivo, toujours délicate en biochimie, devient périlleuse lorsqu'il s'agit de la biochimie de l'oxygène. Rappelons le sort de la réaction de Fenton et des équations d'Haber-Weiss, que nous avons examinées au chapitre IX . Nous verrons au chapitre XV l'évolution du concept de protection par les antioxydants stœchiométriques.
1. O2 : accepteur d'électrons
Le rôle majeur dévolu à l'oxygène au cours de la vie aérobie est d'être l'accepteur final des électrons libérés au cours des divers processus cataboliques, notamment dans la chaîne respiratoire.
La réduction de l'oxygène en eau s'accompagne d'une libération de calories qui est à la source de la plupart des besoins énergétiques des êtres vivants.
Cette réduction peut être accomplie :a) de manière quasi-instantanée au cours de la respiration (e- symbolise l'électron)
O2 + 4 H+ + 4 e- -----> 2 H2O
surtout grâce à la cytochrome oxydase, mais aussi à certaines enzymes cupriques bactériennes et végétales.
C'est dans la mitochondrie que s'accomplit ce remarquable phénomène, principal fournisseur d'énergie de la cellule, que constitue la respiration cellulaire. Les électrons proviennent essentiellement de l'activité de la NADH deshydrogénase et de la succino-deshydrogénase (voir le traité de Lehninger et al., 1993).Un mécanisme évitant la production de ROS
C'est au niveau de la cytochrome oxydase que le prodige a lieu : l'oxygène est réduit instantanément en eau, sans passer par des états réduits intermédiaires (anion superoxyde, peroxyde d'hydrogène : voir chapitre VII), générateurs d'espèces oxygénées activées nuisibles. Toutefois, cette réduction instantanée peut être perturbée au cours de phases anormales du métabolisme. L'hypoxie tissulaire, suivie de ré-oxygénation, entraîne une réduction mono-électronique d'O2 en amont de la cytochrome oxydase (question qui sera traitée dans le tome II, L'oxygène en pathologie des mammifères).
Les problèmes suscités par l'instantanéité de cette réduction totale de la molécule d'O2, sans étapes discernables, au niveau de la cytochrome oxydase sont loin d'être résolus.
On est loin de la vue simpliste des spécialistes de la chaîne respiratoire pour qui l'oxygène n'était qu'une poubelle à électrons recevant ceux-ci sans autre complication.b) de manière progressive
Par les réductions à 1 ou 2 électrons mettant en jeu diverses enzymes : NADPH oxydases, NO synthases, xanthine oxydase, etc… avec intermédiaires chimiquement actifs (voir chapitre XIII) :
O2+ 1 e- ----> O2• anion superoxyde, étape endergonique (qui exige l'intervention de catalyseurs ou d'oxydases).
La dismutation de O2• est considérablement accélérée in vivo par la présence de superoxyde dismutases (chapitre XV).O2• + 2 e- + 2 H+ -----> H2O2 peroxyde d'hydrogène ou eau oxygénée : in vivo, la réduction de H2O2 peut être spontanée, mais étant plus lente, elle permet à des catalyseurs intra-cellulaires comme les ions oxo-ferryles d'intervenir en produisant de dangereuses espèces oxygénées activées.
Dans les cellules à métabolisme élevé, on trouve la catalase et dans la plupart des cellules la glutathion peroxydase. Ces deux enzymes accélèrent considérablement la décomposition de H2O2; elles sont décrites dans le chapitre XV.
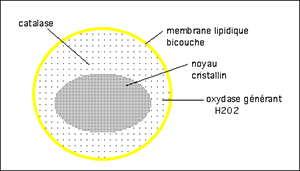
Le tissu hépatique est riche en catalase qui est localisée dans des organelles, les peroxysomes (fig. XII-1). Ceux-ci sont limités par une membrane simple, sphérique, et sont le siège d'un intense métabolisme: synthèse des acides biliaires, du cholestérol, du plasmalogène, mais aussi du catabolisme des purines (xanthine-oxydase), ainsi que les β- et ω-oxydations des acides gras à longues chaînes. Le catabolisme oxydatif intense des peroxysomes explique la présence de catalase dans ces organelles.
Fig. XII-1 : Peroxysome dans une cellule hépatique, examinée en microscopie électronique.
2. O2 : synthèse de molécules physiologiquement actives
Diverses molécules intervenant dans le contrôle des fibres lisses et par là dans l'activité respiratoire (régulation du diamètre des bronchioles) et dans la circulation sanguine (vasoconstriction ou vasodilatation) dépendent directement de l'oxygène moléculaire, car leur synthèse est effectuée par des oxygénases. Notons : NO•
Nous avons vu les propriétés radicalaires du monoxyde d'azote (chapitre VII). Nous verrons dans le chapitre XIII les particularités des NO• synthases qui sont des oxygénases.
Le NO• possède des propriétés vasodilatatrices et bronchodilatatrices remarquables (EDRF de la fin de la décennie 1970). Il intervient dans la neurotransmission.Prostaglandines
Les prostaglandines sont des composés cycliques formés à partir d'acides gras poly-insaturés à chaîne de 20 carbones dont le type est l'acide arachidonique. Elles interviennent sur la contractilité des fibres musculaires lisses des vaisseaux et de l'utérus. Les thromboxanes dérivent des prostaglandines et interviennent dans l'agrégation des plaquettes et, par cette voie, dans la coagulation sanguine. Les prostaglandines font partie des messagers de l'inflammation (chapitre XIII). Nous verrons dans le chapitre XIII les particularités des PG synthases qui sont des dioxygénases hémiques.Leucotriènes
Ce sont des composés aliphatiques dérivant, comme les prostaglandines, d'acides gras poly-insaturés à 20 carbones. Ils sont synthétisés par des lipoxygénases, enzymes à fer non hémique (voir chapitre XIII).
3. O2 : générateur de ROS contrôlant les fonctions cellulaires et la transduction du signal
Il s'agit là d'un domaine de recherches en pleine évolution. Les premiers travaux sont probablement ceux de Mittal et Murad, en 1977, qui attribuèrent aux radicaux formés en aval de la cascade de l'anion superoxyde la stimulation de la guanylate cyclase, productrice de ce que l'on nomma à l'époque le second messager, le cGMP. Plus tard, le rôle primordial de NO• fut mis en évidence. Les travaux actuels montrent que H2O2 joue un rôle indirect dans l'expression de divers gènes en produisant des médiateurs qui sont probablement des ROS. Le lecteur trouvera dans une monumentale revue écrite par W. Droge, en 2002, un panorama approfondi de la question.4. Bibliographie
Pour la respiration mitochondriale :
Lehninger AL, Nelson DL, Cox MM. Principles of Biochemistry. Worth Publ. ed., 1993, pp. 542-597.Pour le rôle des ROS dans la transmission des signaux intra-cellulaires :
Mittal CK, Murad F. Activation of guanylate cyclase by superoxide dismutase and hydroxyl radical: a physiological regulator of guanosine 3',5'-monophosphate formation. Proceedings of the National Academy of Sciences U S A. 1977; 74: 4360-4.
Droge W. Free radicals in the physiological control of cell function. Physiological Review. 2002; 82: 47-95.